|
Inverse Correlation Between Extrinsic Mortality Rate and Proportion of Deaths Due to Intrinsic Mortality |
|
|
Inverse Correlation Between Extrinsic Mortality Rate and Proportion of Deaths Due to Intrinsic Mortality |
From the observational data about mammal and bird life tables in the wild reported and analyzed in an authoritative and documented review [Ricklefs 1998], a very interesting relation was described.
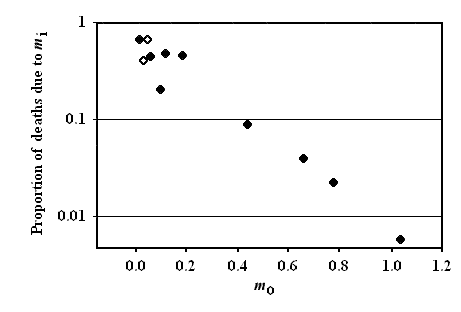
As a matter of fact, data show a significant (p < 0.001) inverse relation between extrinsic mortality (m0) and the proportion of deaths due to intrinsic mortality (mi) (Fig. 1).
FIGURE 1 – Inverse significant correlation between m0 and the proportion of deaths due to mi. Data are from [Ricklefs 1998], Table 2 (p. 30). Ricklefs’ fig. 7 (p. 34) has been redrawn. Ordinates are in logarithmic scale. Open symbols refer to mammal species, solid symbols to bird species.For non-adaptive hypothesis as Wear and Tear and Stochastic Hypotheses, Mutation Accumulation Hypothesis, Antagonistic Pleiotropic Hypothesis and Disposable Soma Hypothesis, there is a clear prediction:
“The principal determinant in the evolution of longevity is predicted to be the level of extrinsic mortality. If this level is high, life expectancy in the wild is short, the force of selection attenuates fast, deleterious gene effects accumulate at earlier ages, and there is little selection for a high level of somatic maintenance. Consequently, the organism is predicted to be short lived even when studied in a protected environment. Conversely, if the level of extrinsic mortality is low, selection is predicted to postpone deleterious gene effects and to direct greater investment in building and maintaining a durable soma” [Kirkwood & Austad 2000].
Ricklefs plainly states in his discussion (not in the title of his paper!) that this prediction is clearly contradicted by the inverse relation observed between m0 and the proportion of deaths due to mi [Ricklefs 1998].
This contradiction has been clearly underlined [Libertini 2008] and so far no one of the supporters of non-adaptive theories has formulated (or tried to formulate) an explanation.
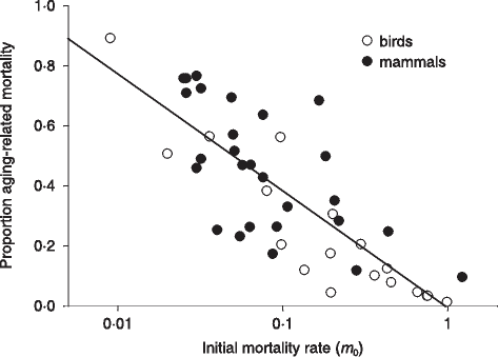
Ricklefs has confirmed his data in a more recent paper [Ricklefs 2008], trying to explain them saying that "because so many individuals suffer ageing-related deaths, it appears that the further evolutionary postponement of senescence is constrained by limited biological potential for prevention and repair of physiological decline". This non-explanation is in clear contradiction with Kirkwood & Austad's statements and with the position of the same Ricklefs in 1998.
FIGURE 2 – The same of fig. 1 but the figure is from [Ricklefs 2008].References:
- Kirkwood, T.B.L. & Austad, S.N. (2000) Why do we age? Nature 408, 233-238. [PubMed] [Google Scholar]
- Libertini G. (2008) Empirical evidence for various evolutionary hypotheses on species demonstrating increasing mortality with increasing chronological age in the wild. TheScientificWorldJOURNAL 8, 182-93 DOI 10.1100/tsw.2008.36. [PubMed] [Google Scholar] [Free]
- Ricklefs, R.E. (1998) Evolutionary Theories of Aging: Confirmation of a Fundamental Prediction, with Implications for the Genetic Basis and Evolution of Life Span. Am. Nat. 152, 24-44. [PubMed] [Google Scholar]
- Ricklefs, R.E. (2008) The evolution of senescence from a comparative perspective. Funct. Ecol. 22, 379-392. [Google Scholar] [Free]
www.Programmed-Aging.org
Sponsored by Azinet LLC © 12/2009