|
Theoretical Arguments Against Mutation Accumulation Hypothesis |
|
|
Theoretical Arguments Against Mutation Accumulation Hypothesis |
Against Mutation Accumulation Hypothesis as possible explanation of aging it has to be remarked that: “As pointed out by Mueller and Rose (1996), at some point selection becomes weaker than the forces of mutation and genetic drift and adaptive evolutionary responses cease. However, this point occurs well after 99% of individuals in a population have died, and the mechanism therefore is not relevant to mechanisms of senescent death occurring up to this point” [Ricklefs 1998]
A possible objection is that Mueller and Rose’s statement could be dubious in the case of the combined action of many harmful genes. In regard to this possibility, a simple theoretical argument was formulated [Libertini 1988].
In a non-aging population the life table of which is given by the equation:
(1) Yt = Y0 × (1 – λ)t
where: Y0 = starting population; Yt = survivors at time t; λ = death-rate, if C is a dominant allele expressing its disadvantageous action (S) only and exclusively at the time t (“t-gene”), C' its neutral allele, V mutation rate of C' in C (while mutation rate of C in C' is considered negligible), then the frequency of C at the (n + l)th generation is given by:
(2) Cn+1 = [Cn × (1 – S × Yt – V) + V] / [1 – Cn × S × Yt]
The equilibrium frequency of C (Ce), namely the frequency of C when Cn+1 = Cn, is given by:
(3) Ce = V / (S × Yt)
and the increment of the mortality rate at the age t (Δmt) will be:
(4) Δmt = Ce · S = V / Yt .
Supposing the existence of n t-genes for each age t, all the genes for sake of simplicity with equal values of V (the value of S is irrelevant), at each age:
(5) Δmt = n · V / Yt ,
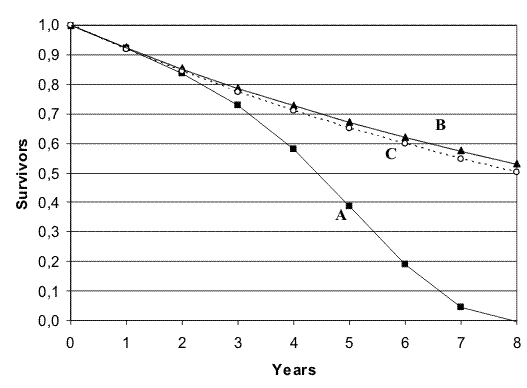
namely Δmt follows the decrement of Yt (=λ), increasing slowly until Yt is very small and only then becoming exponential.In Fig. 1, curve A is the life table of Rangifer tarandus (M) (for the simulation, Weibull’s equation, mt = m0 + α × tβ, has been used, with m0=0,076, α=0,00203; β=2,9968; equation and data are from Ricklefs [Ricklefs 1998] (see p. 27 and table A2 in Appendix A -); curve B is a hypothetical life table of the same species with only the extrinsic mortality at its lowest value (m0); curve C is a hypothetical life table with m0 plus the effects of a great number of t-genes with high mutation rates from their inactive alleles (n = 500; V = 0.00001). Notwithstanding the high values of n, curve C is quite different from curve A and the area between curve A and curve C is completely unjustifiable as effect of t-genes insufficiently eliminated by selection.
The same argument with appropriate modifications can be put forward for recessive t-genes too. In such a case:
(3') Ce = SQR(V / (S · Yt))
and
(4') Δmt = Ce2 · S = V / Yt ,
as for dominant t-genes.
FIGURE 1 - Effect of many t-genes on a life table (for explanations, see text).
In short, even with extreme conditions the decline of Yt in the wild, namely aging, cannot be the consequence of insufficient selection against t-genes and Mutation Accumulation Hypothesis cannot be proposed as an explanation of aging.
References:
- Ricklefs, R.E. (1998) Evolutionary Theories of Aging: Confirmation of a Fundamental Prediction, with Implications for the Genetic Basis and Evolution of Life Span. Am. Nat. 152, 24-44. [PubMed] [Google Scholar]
- Libertini, G. (1988) An adaptive theory of the increasing mortality with increasing chronological age in populations in the wild. J. Theor. Biol. 132, 145-162. [PubMed] [Google Scholar] [Free]
www.programmed-aging.org
Sponsored by Azinet LLC © 2009